有關細節參見細節參見黃色字體浮框或文章:: Quantum Gravity Non-simultaneous EP Irrotational Entropy Preservation Origin of Life
Download: http://doi.org/10.21275/SR251231101432
北京醫諾安醫藥科技有限公司
基礎研究捐款-應用研究投資公賬:
中國建設銀行清華大學支行:
1105 0163 6000 0000 0742
有關捐款或投資事宜請聯繫:Lab@innoen.org
聯繫我們:lab@innoen.org, webchat
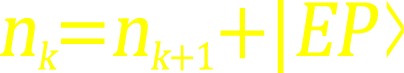
奇點(singularity)通常指在理論計算中,某些物理量趨於無窮大、不可定義,或原本應彼此獨立的多個物理量被迫在同一點同時坍縮的情形。例如,引力時空奇點即是廣義相對論框架下,由幾何坍縮所產生的極限點。從更直觀的角度看,在傳統幾何體系中,常常默認可以將(x+∣x'⟩, y+∣y'⟩)絕對性地簡化處理為(x,y), 即認為不確定項∣x'⟩,∣y'⟩可以無條件被幾何歸零(zerobility). 這種約定俗成處理在牛頓力學框架下多數可近似接受, 但是當這種絕對歸零的處理方式不再成立時,便會遭遇瓶頸,並以“奇點”的形式表現出來。因此,奇點並非自然本身的物理屬性,而是舊幾何體系固守絕對歸零性(absolute zerobility)所產生的結構性缺陷.  帥在今天的漢語裏面仍然保留有帥氣,英俊,瀟灑等意思,所以“氣為血之帥,血為氣之母”有三層內涵:第一,氣為血的父親,第二,氣是父親中最優秀的父親,並非普通的父親.第三,最優秀父親仍然是由母親所生下,这样下来氣和血的關係就由“帥”和“母”完全界定.用西方醫學概念,氣是先天免疫,例如基因層面的CRISP/CAS9,而血就是血液後天獲得性免疫,如T細胞,B細胞等等.對於人體來説,先天免疫只能通過合理的spinal converging物理訓練逐步積累,完全無法通過藥物或注射获取,後天免疫則可以在一定程度上用現代藥物或注射去彌補.Long COVID-19危機就是不良藥企妄圖用注射方式強化人體先天性免疫失敗的典型案例. 此外.所謂類似抗腫瘤疫苗之類也是難以實現的故事,因為對抗腫瘤只能靠先天物理免疫原性而無法靠後天藥物或注射產生的獲得性免疫. 妄圖用後天獲得性免疫方法,例如疫苗,CAR-T編程再注射,干细胞等等蠱惑大眾付費去對抗腫瘤,實際是一種商業利潤操作而非醫學, 這種商業故事是無法導致醫療上的抗癌奇蹟出現的. 

跪盤(伸筋強肝延壽盤),是中國古代橫跨商朝(源於商代1300BCE權貴階層專用,到秦漢221BCE擴展為全民禮儀必用)直到到清滅亡(-1912 CE)的古代中國全民抗逆生活約束方式.該法在今天的日本仍有文化性留存.这种方法確實是全世界最高水平的俗家先天免疫能力物理訓練方法,若自幼长期以此姿态替代常规座凳,即使不進行任何傳統功夫或現代體育訓練,也不刻意改变日常生活方式,都能在同等條件下延壽十年以上,應該算是中華民族對世界健康事業貢獻最偉大的瑰寶.該俗家方法的功家升級版為雙盤,而金剛鐵板橋作為全世界最高水平的專業功家先天免疫抗逆訓練則僅在中國武當太和門流傳.該法把少林恨天無把(懸脖功)加上武當恨地無環(懸腳功)最後用金剛鐵板橋搭橋上架,歷時數十年方成.君穴對為肚臍(元氣眼)與頭頂发漩(百會)在最高抗逆的橋架下實現脊柱CSF熵留存:nk=nk+1+∣EP⟩,去驱動其它“臣”,“佐”,“使” 等一系列次級穴位實現熵留存.所有中醫穴位都處於活體不對稱自旋(asymmetric spin) 狀態才能有效獲取∣EP⟩.這裏我們可以把人體穴位看成多陀螺儀體系,人體最大先天陀螺儀的兩錨點就是君穴百會和肚臍,irrotational entropy驅動力使得這对陀螺儀锚点帶動其它各級別穴位陀螺儀spin,從而指向自己正確方向.注意,所謂先天免疫就是這些陀螺儀功能強勁指向正確而所謂衰老就是這些陀螺儀旋轉的系統inversion衰减而導致指向紊亂.中醫醫家穴位,武家穴道穴盤等的無旋熵流,只存在於活體,無法進行解刨學觀察.需要注意的是,無論一個飛行器上裝有多少個陀螺儀,都會指向同一個方向,但是人體針灸穴位先天免疫陀螺儀或者四極子(串),所有指向必定不同,因為它們之間並不是“薛定諤貓”式的zeroable經典量子疊加,而是∣EP⟩參與的 non-zeroable inversion疊加而積累irrotationality, 不可歸零性(non-zeroability) 培育了各向異性穴位spin慣性.只有當生物體死亡以後,這個多陀螺儀或quadrupole系統才會最終塌陷(or zeroable)成同一方向.(Ref:自旋的熵穩定性重心.在牛頓力學和量子力學下,自旋都是重力無關參數,僅僅在相對論框架下,存在一種極其微弱的耦合.然而,在生物系統中,與自旋相關的引力效應可以通過旋轉動量被指數級放大.由此可定義一種熵穩定性“重心”.在傳統量子力學中,微觀對稱自旋通過旋轉對稱性的表示論被併入角動量守恆框架(𝐽=𝐿+𝑆),從而在形式上被歸入能量守恆的範疇.然而,這種定義上的關聯主要是代數層面的,而非基於明確物理機制的結果.自旋所體現的穩定性,更自然地應歸屬於熵留存(entropy preservation)的範疇.按照新的理論體系,若缺乏 EP作為橋接,自旋無法與能量守恆參數共同置於同一物理方程中.)武當太和門穴位系統(無旋熵引力穩定性“重心”)跟醫家書籍上的不太一樣,分三十六天罡和七十二地煞,還進一步演化出範圍更大的穴道,穴盤等.目前市面上少林功夫書籍所發表的只有沿用中醫書上的穴位,或許是因為戰亂文革等原因,多數少林七十二藝功夫原本已經失傳,這類沿用中醫書上穴位的少林功法出版物,更像是後人補作,不太像是少林嫡傳原貌 (因為穴位限於醫家針灸所用,功家所用的應該包含更大範圍的穴道和穴盤).  

熱力学熵是由卡诺循环歸遞出來,按照这一体系,不可逆循环导致熵增ΔS≥Q/T ,但是当熵增能满足生物量子化条件后,所有in vivo过程就不再按薛定谔猫方式叠加而改为nk=nk+1+∣EP⟩
方式叠加,其中nk, nk+1等是整数,围绕∣EP⟩糾纏積累non-simultaneous time,生命有限就因為生物體只能存活在“不歸零同時性時間(non-simultaneous)”狀態下而无法使用rigid time. 我們可以簡單的説, non-simultaneous time 就是rigid time通過表面張力吸收EP以後的表達形式.香農熵(Shannon Entropy)是一種用uncertainty定義的熵, 傳統的rigid time之穩定性是由牛頓公理的絕對歸零型(absolute zeroability) 所預先贈予的, non-simultaneous time所需要的穩定性,則是從香农熵位(Shannon Entropy Loci)得到的.無論這類non-simultaneous time怎麼變化,必須依賴其香農熵位獲取足夠穩定性. 也就是説玻爾茲曼的chaotic entropy是缺乏足夠穩定性的熵,而irrotational entropy 則是靠香農熵位賦予的穩定性而實現量子化的熵 (注意:熱力學第二定律只對缺乏香農熵位穩定性的chaotic entropy有效, 對具備由重力提供香農熵位穩定性的rotational entropy失效,原因是這個定律本質上是剛體參數模式而從未觸及非剛體範式). 傳統量子力學波函數以及自旋spin跟重力或熵毫無關係.但是生物量子態必須依賴重力提供的熵穩定性才能實現量子化,從而生長和進化.
Non-simultaneous time是具備香農熵位穩定性的時間,rigid time不具備香農熵穩定性,只包涵牛頓座標穩定性或愛因斯坦彎曲空間曲率穩定性.同理,經典量子力學的對稱性spin是不具備熵穩定性重心的,只有不對稱spin才具備熵穩定性重心.這裏涉及到重心這個牛頓時代的名詞之時我們需要提示一下,牛頓的萬有引力定理由rigid重心所測得的引力常數GCavendish,遠遠低於生物活體由無旋熵重心所表達的引力常數Gbio-system.G實驗偏差如果按Cavendish的方法計算可以高達109數量級差距,也就是说熵和時間都有不可歸零的“重量”或“重心”.注意:這種不歸零性(non-zeroability)無法被歐幾里得幾何或傳統非歐幾何完整描述;即便是愛因斯坦建立廣義相對論所依仗的黎曼幾何,在本質上仍然是建立在可局域歸零的幾何假設之上而無法對此完整刻畫.相反,音樂Chu’s constant居然可對此進行比較好的描述.  根據Landau的極低溫凝聚態二流體模型,超流體是一種不再消耗體系能量的狀態,由此延伸出的熵留存定律nk=nk+1+∣EP⟩描述了超流體部分,常流體部分以及環境之間的關係.這種理論延伸需要把傳統內稟屬性基础微观對稱性自旋(無衰老週期)升級到表面張力基础(遵循衰老週期)的可宏观延申不對稱自旋.也只有延伸後的不對稱自旋體系才能用於生物體系由∣EP⟩主導的遺傳和進化,所以生物體系也可以稱為常溫(∣EP⟩無旋熵宏觀)凝聚態實體(生物通過表面張力而非低溫達到condense).就人體而言,各種in vivo關係,無論宏觀的,如眼看到,耳聽到,鼻聞到,口嚐到的各種信息;人體各個器官組織,肌肉韌帶等之間的關係;還是微觀的,比如各種DNA/RNA,蛋白質還是各種細胞之間的關係,只有一種無旋熵紐帶,也就是説上述所有關係都能用熵簡併定律nk=nk+1+∣EP⟩的各種不同水平進行描述,生物體內部結構中並不存在任何超出這個定律之外的關係.在傳統生物學中,視覺和聽覺的描述差別很大,人們很難找到這兩鍾感官之間的關係,但是熵留存定律改變拉這種盲人摸象的局面.傳統量子力學只能用於微觀世界,量子物理學家自己都不認為他們的工作跟生物學研究有任何關係,但是現在有了這個熵留存定律nk=nk+1+∣EP⟩,所有生物無論大小和進化程度高低,只要是活體,一定能用到這個定律進行量子化描述,生物脊柱就是一個肉眼可見的量子化基本結構.現代分子生物學與遺傳學所建立的基因組學、蛋白組學以及各類代謝關係和相互作用框架,大多成型於以能量守恆為核心範式的時代.隨着研究對象和理論邊界的變化,這些體系中的相當一部分有必要進行實質性的修訂,以適應以熵留存(entropy preservation)為核心的新範疇.生物不對稱自旋的熵穩定性“重心”源於表面張力,傳統量子力學由於各位先驅們省略了表面張力和重力影響,只剩下對稱自旋作為唯一選項.這種僅剩的選擇也使得其自旋不得不關聯於能量守恆範疇從而不可避免的屏蔽了其entropy preservation的本質特徵.特別注意的是,對稱自旋(symmetric spin)與非對稱自旋(asymmetric spin)有五點数学物理技巧上的不同:第一,发生条件不同,對稱spin是一種可以在微觀任意位置出現的內稟屬性,而不對稱spin只能發生在超流體部分(在旋轉動量matrix中,超流體部分和常流體部分不可分,可延申到宏觀世界發生);第二,動力學響應機制不同,對稱spin具有Bohr spooky特徵,对纠缠态中的一个自旋进行测量,另一个自旋将瞬时塌缩到相反方向,其塌縮表觀速度遠超光速.相比之下,在非對稱spin中,诸如nk及nk+1等態之間並不存在這種“超自然”的瞬時塌縮,實際響應速度遠遠低於光速(以呼吸為例,呼吸過程中肺泡超流體nk及nk+1吸收∣EP⟩進行轉化的速度,最多僅能达達到呼吸節律的數十倍速度而已);第三,與環境的關聯不同,對稱spin是內稟屬性而跟環境無關,但是非對稱spin需要∣EP⟩來維持. 唯有不對稱結構才能讓∣EP⟩攝入,這也是生物只能利用無旋熵維生而無法利用傳統Noether對稱性能量的根本原因.第四,塌縮trigger條件和結果不同,對稱spin可以僅僅因為檢測就collapse到非生命的本徵態,不對稱spin關聯無論在什麼水平一直存在到該水平結構死亡(心臟跳動例子,除非生物體死亡心臟不會停止跳動).這種區别很大程度源於塌縮結果不同,對稱spin塌縮結果是無生物活性的eigenstate,而不對稱spin則是在有生物活性的nk與nk+1之間互跳,只要有∣EP⟩可用就能維繫具備生物活性的linearity shifting.可以説,微观对称spin是無生物活性measurement-induced collapse,而宏观不对称spin具備生物活性structurally persistent aging. 第五,也是最令人費解的一點,我曾向許多物理學家、其中包括我的相對論啓蒙老師提出過問題——既然單質原子氣態下的電子自旋被認為是對稱的,那麼當兩個元素形成化合物之後,例如最簡單的CO₂,甚至像單質的O₂或臭氧分子(O₃),電子自旋是否仍然保持對稱或依然具備Bohr Spook Collapse?大多數人的回答往往是回避,認為那屬於化學家的研究範疇,與物理學無關而拒絕回答;少數嘗試正面回應的人,則多以軌道雜化或相關模型解釋,但這些難以令人信服.既然量子力學引入自旋量子數是為了使理論保持邏輯上的自洽,那麼CO₂ 這樣小分子中電子自旋,為何就不能是C和O中軌道互跳呢,如nk carbon=nk+1_oxygen+∣EP⟩?這第五點應該是量子學科發展瓶頸問題而跟前面四點數學物理技巧不同,但是因為涉及經典量子力學向生物体系靠攏最重要的關鍵所在,所以專門在此提出來. |



|